Введение
Oдним из наиболее важных направлений биологии является, безусловно, эволюционная теория. Более того, выяснение филогенетических связей и эволюционных механизмов должно быть основной задачей фундаментальной биологии, в противовес практической биологии, преследующей строго утилитарные цели наподобие решения экологических, медицинских и продовольственных проблем. Наиболее интересными в трактовке надвидовой эволюции можно назвать, пожалуй, концепции ортогенетического толка, в разработке которых принимали участие такие учёные как Л. Берг, Н. Вавилов, А. Любищев, С. Мейен [Московский, 99], но они, обнаруживая ряд важных закономерностей в эволюционном процессе, придают мало внимания естественным механизмам эволюции. К сожалению, дать полный обзор эволюционных гипотез в работе, не касающейся напрямую их сравнением, нельзя, поэтому интересующимся можно порекомендовать исследование Назарова [Назаров, 91]. Серьёзного внимания заслуживает также концепция Любищева о систематике как «окне в онтологию», согласно которой система живых организмов является не чем-то условным, а изначально «запрограммированной», заложенной в природу системой, наподобие системы элементарных частиц или химических элементов [Московский, 99].
В данной работе представлена попытка оформления умозрительных методов (практически потерявших свою роль в естественных науках со времён натурфилософии, но, тем не менее, актуальных и по сей день) и их применения к эволюционной систематике и теории эволюции как наиболее важным дисциплинам теоретической биологии. В целом работа носит не столько общебиологический, сколько натурфилософский характер, хотя основывается в значительной степени на биологическим материале и предназначена для реализации биологических задач.
Целями данной работы является доказательство актуальности и жизнеспособности умозрительных методов в эволюционной систематике и теории эволюции и выявление с помощью умозрительного анализа основных механизмов эволюции. Для осуществления этих целей поставлены следующие задачи, решаемые в трёх разделах работы:
- Определение, формулировка и систематизация основных принципов умозрительного метода в эволюционной систематике.
- Попытка сформулировать наиболее полное и достаточное определение жизни и, на его основе, попытка представить в общих чертах процесс возникновения жизни на Земле.
- Выявление возможных механизмов эволюции и разделение макроэволюционного процесса на мезо- и мегаэволюцию.
Поскольку эволюционная наука является наиболее важной для теоретической биологии, а многие её вопросы до сих пор являются дискуссионными, имеется огромное количество литературы, в той или иной степени затрагивающие вопросы эволюции и систематики, как в целом, так и по отдельным группам. Данная работа, хотя и основывается на наиболее древних, умозрительных методах научного познания, может рассматриваться как новый подход к общетеоретическим проблемам биологии, поэтому даже в том случае, если отдельные выводы, сделанные в ходе этой работы, будут опровергнуты, умелое применение умозрительного анализа может дать заметный результат для развития науки в целом. Работа предназначена не столько для профессиональных биологов, сколько для простых читателей, пытающих разобраться в вопросах теории эволюции. Многие факты, затронутые здесь, могут оказаться общеизвестными, однако их систематизация и сведение воедино сами по себе должны представлять интерес.
- Умозрительный метод
Kак и любой другой, умозрительный метод имеет свои плюсы и минусы. К плюсам относится его универсальная применимость к общетеоретическим вопросам (то есть, одни и те же принципы могут применяться при построении гипотез в физике, астрономии, истории, теологии, биологии и т. д.), отсутствие абсолютной необходимости конкретных узкоспециальных знаний и навыков (хотя и они, разумеется, весьма желательны), практическая простота (это не значит, однако, что умозрительный метод прост и общедоступен: просто, если человек владеет им, ему не нужны ни оборудование, ни строго заданные условия проведения исследований); наконец, основным плюсом метода является то, что он позволяет сводить уже имеющиеся факты в единую картину. С другой стороны, метод этот бесполезен в практических отраслях науки, не предназначен для прямой, механической передачи другим людям, и его результаты могут не согласовываться с какими-либо фактами, неизвестными исследователю или не принятые им в расчёт; однако метод обладает особой пластичностью, и при выявлении новых фактов его результаты могут легко изменяться этим или другим исследователем без существенных изменений в самой сути гипотезы.
Основной задачей метода является анализ и синтез наибольшего числа фактов и концепций (если последние, разумеется, не находятся в непреодолимом противоречии друг с другом), а не подтверждение или опровержение той или иной конкретной гипотезы. Значение умозрительного (гипотетико-дедуктивного и индуктивного) метода подчёркивается и в работе Назарова [Назаров, 91, стр. 24]. Так, очевидно, что такие важные естественнонаучные методы как наблюдение и эксперимент слабо применимы к надвидовой эволюции (макроэволюции в традиционном понимании). Исходя из этого, единственный верный путь для всестороннего изучения макроэволюции — реконструкция её механизма и построение соответствующей гипотезы на основе сопоставления и объединения данных, относящихся к разным уровням биологической организации. При этом важнейшим критерием достоверности умозрительной гипотезы является «требование взаимной логической и фактической непротиворечивости данных» [Назаров, 91, стр. 47-48]. Именно таким положением мы руководствовались, описывая механизмы и результаты эволюции на надвидовом уровне.
1.1. Классификация принципов умозрительного метода
Принципами умозрительного метода в эволюционной систематике и теории эволюции (ПУМ) названы здесь некоторые достаточно разрозненные и пока что слабо систематизированные тезисы, касающиеся закономерностей эволюционного процесса, биологической систематизации и действия самого метода, которые следует учитывать при умозрительном анализе теории эволюции и эволюционной систематики для построения наиболее чёткой и логически выверенной эволюционной картины. Для начала необходимо отметить некоторые из них (как сформулированные, так и не сформулированные ранее).
ПУМ можно разделить на три основные группы: принципы действия метода (ПДМ), принципы определения (ПО, подразделяющиеся напринципы возможности, возникновения и направленности) и принципы систематизации (ПС) (см. табл. 1) . Следует отметить, что, во-первых, такое разграничение описанных ПУМ достаточно условно, так как один и тот же принцип может иметь разные аспекты и применяться к разным ситуациям, а, во-вторых, что перечень принципов, вероятно, можно расширить. Не исключено также, что в дальнейшем, при более подробной разработке синтетической теории эволюции, некоторые из перечисленных здесь принципов будут увязаны с положениями о механизмах и движущих силах эволюции и войдут в эволюционную теорию и систематику, а также в общебиологическую методологию.
Таблица 1. Принципы умозрительного метода
| Принципы умозрительногометода | 1. Принципыдействия
метода |
1.1. Принципмозаичности | |
| 1.2. Принциппрезумпции
позитивизма |
|||
| 1.3. Принциписключений | |||
| 1.4. Принциплабильности |
| 2. Принципыопределения | 2.1. Принципывозможности | 2.1.1. Принципвероятности
|
|
| 2.1.2. Принципкомбинирования | |||
| 2.2. Принципывозникновения | 2.2.1. Принциппарафилетичности | ||
| 2.2.2. Принципвариабельности | |||
| 2.2.3. Принцип тупико-вости специализации | |||
| 2.2.4. Принцип фило-генетического ускорения | |||
| 2.3. Принципынаправления | 2.3.1. Принципэкономичности развития | ||
| 2.3.2. Принципизъятия лишнего | |||
| 2.3.3. Принцип экологи-ческого давления | |||
| 2.3.4. Принцип эволюци-онных тенденций | |||
| 2.3.5. Принцип холизма | |||
| 2.3.6. Принципнаправленности | |||
| 2.3.7. Принципдопустимости | |||
| 2.3.8. Принципсопряжённости | |||
| 3. Принципысистематизации | 3.1. Принципдетерминирования | ||
| 3.2. Принцип кладизма | |||
| 3.3. Принцип унификации | |||
| 3.4. Принцип повышениястатуса |
1.2. Принципы действия метода
К принципам действия метода относятся принципы мозаичности, презумпции позитивизма, исключений и лабильности.
Принцип мозаичности. Любая полнота фактического материала недостаточна для полного прослеживания эволюционных связей.Исходя из этого принципа, вне зависимости от имеющихся данных (прежде всего палеонтологических, так как многие эволюционные преобразования могли не оставить никакого следа в палеонтологической летописи, и биохимических, так как с точки зрения теории вероятности выявить все биохимические особенности всех ныне живущих и тем более вымерших видов невозможно) для эволюциониста-систематика необходимо умение создавать умозрительные построения.
Принцип презумпции позитивизма таков: если какое-либо утверждение не опровергнуто и не доказано, то его следует считать возможным. Исходя из этого принципа, все механизмы, которые когда-либо предполагались в качестве движущих сил эволюции и не были достоверно опровергнуты, можно допускать как имевшие или имеющие место в эволюционном процессе.
Принцип исключений можно афористически сформулировать следующим образом: если закон подтверждается исключениями, то он плохо сформулирован. Другими словами, всякое имеющее место исключение должно восприниматься не как подтверждение правила, а именно как исключение, и появление каждого нового исключения из старого правила, каким бы незначительным оно ни казалось, требует изменения правила.
Наконец, важен в умозрительных построениях принцип лабильности, согласно которому всякие новые данные могут быть уложены в прежнее построение умозрительным же способом.
1.3. Принципы определения
Принципы определения подразделяются на принципы возможности, принципы возникновения и принципы направления в зависимости от того, какую часть эволюционного процесса они затрагивают.
К принципам возможности относятся те ПУМ, которые рассматривают вероятности появления тех или иных признаков у разных групп живых организмов. Нами сформулированы два принципа этой группы: принцип вероятности и принцип комбинирования.
Согласно принципу вероятности, для установления родственных связей между видами и группами, обладающими сходными структурными, экологическими, физиологическими и другими признаками, следует выяснить, насколько вероятно их моно- или полифилетическое возникновение. При этом очевидно, что чем сложнее признак, чем из большего числа составляющих он состоит (или же чем больше сходных признаков у данных видов), тем больше вероятность, что группа, им обладающая, является монофилетической (в широком смысле слова, включающем также понятие «парафилетический»); обратная формулировка при этом не верна: простой признак может возникнуть как поли-, так и монофилетически. Так, высока вероятность того, что жгутик эукариот, обладающий сложной упорядоченной структурой 9+Х (где Х может быть равно 0, 1 или 2), возник монофилетически (в данном случае не рассматривается возможность его полифилетичного приобретения путём симбиоза согласно гипотезе Линн Маргелис: возникла эта структура, в любом случае, скорее всего, монофилетично, пусть и у гипотетических прокариотических симбионтов [Маргелис, 83]), тогда как простой флагеллиновый жгутик прокариот мог возникнуть как один, так и несколько раз независимо.
Принцип комбинирования гласит, что если какой-то признак мог возникнуть в одной группе, то существует ненулевая вероятность его возникновения в другой, причём, с одной стороны (исходя из принципа вероятности), чем сложнее признак, тем эта вероятность меньше, а с другой (исходя из принципа эволюционных тенденций) — чем группы эволюционно ближе друг к другу, тем она больше. Из этого ПУМ также следует, что возможно существование «химерной» группы, сочетающей признаки других групп, но вероятность существования такой группы уменьшается при увеличении числа и сложности этих признаков, а также при удалённости друг от друга и от гипотетической «химеры» реальных носителей этих признаков. Принцип этот следует учитывать при умозрительных эволюционных построениях, однако следует помнить, что он значительно теряет свою применимость в случаях симбиогенетического происхождения признака и приобретения признака путём горизонтального переноса генов.
Принципами возникновения называются ПУМ, оперирующие с базовыми группами, в которых происходит возникновение и накопление признаков. К ним относятся принципы парафилетичности, вариабельности, тупиковости специализации и филогенетического ускорения.
Согласно принципу парафилетичности, монофилетичность примитивного признака не свидетельствует о монофилетичности прогрессивного признака того же эволюционного ряда. Чем древнее признак, тем больше вероятность его монофилетичности: группа, обладающая примитивным признаком, скорее всего, является базовой, парафилетической, а от неё моно- или полифилетично (причём под монофилией может подразумеваться не только происхождение от единственной особи или даже популяции, но и от вида в целом, при использовании идентичных путей видообразования в разных популяциях) отделяются группы с одинаковыми или различными прогрессивными признаками. Примером здесь может являться конечность млекопитающих: млекопитающие с пятипалой стопоходящей конечностью составляют парафилетический пласт, от которого одним или разными путями отпочковываются ветви, ведущие к копытному пальцехождению через некопытное пальцехождение (к конечности, подобной лошадиной, через конечность, подобную собачьей; причём, вероятнее всего, такая эволюция полифилетично привела к возникновению разных групп копытных), а также к многочисленным другим модификациям лапы [Наумов, Карташёв,79, стр. 207-208].
Принцип вариабельности заключается в том, что в древних группах признаки могут быть не устоявшимися, то есть даже близкородственные виды могут иметь существенные различия, на более высоких этажах эволюции считающиеся систематически значимыми. Так, у кишечнополостных обнаружены все основные типы дробления: от примитивного хаотического до примитивных форм спирального и радиального[Догель, 80; Захваткин, 49], — тогда как у более прогрессивных групп характер дробления лежит в основе разделения на две большие ветви —Spiralia и Radialia [Адрианов, Малахов, 85]. Это говорит о том, что в примитивных группах может формироваться определённый фонд для дальнейших преобразований, из которого берут начало прогрессивные ветви.
С принципом вариабельности связан также принцип тупиковости специализации. Согласно этому принципу, чем более специализирован организм, тем меньше вероятность его эволюционирования (кроме микроэволюции, не зависящей от степени специализации [Грант, 80, 91]). Действительно, вид, специализирующийся, допустим, на строго определённой пище (стенотроф), гибнет при её отсутствии, а не перестраивается на другую пищу; крыло летучей мыши, копыто оленя или ласт моржа также не может эволюционировать ни во что другое, на что любят напирать сторонники креационизма [Жизнь — как она возникла? 85]. Этот ПУМ является одним из наиболее важных для осознания изложенных ниже представлений о механизмах эволюционного процесса, так как, с традиционной точки зрения, полностью противоречит самому факту эволюции, поскольку большинство современных видов, если не все, в чём-либо специализированы. Однако это отнюдь не свидетельствует о том, что эволюция в данный момент прекратилась или не шла вовсе: просто подходить к ней нужно с совершенно других позиций (а именно — с учётом роли неотении). Для разъяснения этого принципа следует отметить также, например, тот факт, что приматы как отряд с биологической точки зрения довольно примитивны, что и дало им возможность для эволюционного скачка в плане развития ЦНС.
Принцип филогенетического ускорения общеизвестен. Он сформулирован Бергом и заключается в том, что «индивидуальное развитие может не только повторять филогению, но и предварять её» [Берг, 77, стр. 88].
Принципы направления (принципы экономичности развития, изъятия лишнего, экологического давления, эволюционных тенденций, холизма, допустимости, направленности и сопряжённости) связаны с механизмами, направляющими эволюцию.
Согласно принципу экономичности развития, эволюция идёт преимущественно по наименее энергоёмкому пути. Из этого принципа вытекает и положение о необратимости эволюции [Кейлоу, 86] (не применимый, однако, к процессам микроэволюции): абсолютно неэкономичен (и потому нереален) выход на определённую эволюционную ступень с последующим возвратом на прежний уровень (упрощение организмов под воздействием прикреплённого или паразитического образа жизни, возврат в водную среду или, например, вторичный переход обезьян-панид к четвероногому способу передвижения от распрямлённых двуногих предков — не эволюционный возврат, а крайние формы прогрессивной эволюции под воздействием специфических условий среды; иными словами, катагенез есть видоизменённая форма арогенеза). Так, нелогично предполагать, что выделяющие мочевину млекопитающие происходят от выделяющих мочевую кислоту рептилий, происходящих, в свою очередь, от выделяющих мочевину амфибий [Наумов, Карташёв, 79, т. 2, стр. 46-47]: гораздо логичнее выглядят предположения о независимом происхождении рептилий и млекопитающих от разных групп амфибий либо даже о происхождении амфибий, рептилий и млекопитающих (или, что вероятнее, различных групп «амфибоидов», а также анапсидных, диапсидных и синапсидных позвоночных) от разных групп рипидистиевых кистепёров (а возможно даже, некоторые группы наземных позвоночных произошли от двоякодышащих или иных групп лопастепёрых [Гангус, 2001]). Примечательно, что к аналогичному выводу приводит нас и сравнение суставов конечностей амниот, где одну из групп образуют современные амфибии и млекопитающие, а другую — рептилии и птицы [Наумов, Карташёв, 79, т. 2, стр. 207].
Близок к принципу экономичности принцип изъятия лишнего, согласно которому вызывающие энергетическую неэкономичность органы, процессы или стадии развития утрачиваются или упрощаются до минимально необходимых. Таким образом, виды, обладающие «балластными» признаками, от которых теоретически можно избавиться без вреда для организма, скорее всего, являются более примитивными, чем те, у которых ничего лишнего нет. На основе этого принципа можно, например, выстроить эволюционный ряд изменения процесса гаструляции:мультиполярная иммиграция — униполярная иммиграция — плотное врастание — инвагинация, — так как движение клеточных пластов более «совершенно» и экономично, нежели несогласованное движение клеток [Беклемишев, 79, стр. 38; Богомолов, 68; Захваткин, 49; Зусман, 73; Иванов, 68].
Принцип экологического давления в какой-то степени противоположен принципу изъятия лишнего и заключается в том, что наиболее долго в неизменном виде сохраняется тот признак, который менее всего испытывает давление окружающей среды. Согласно этому принципу, наиболее долго сохраняются именно признаки, не играющие существенной адаптивной роли, однако признаки эти обычно мало обращают на себя внимание. Хорошим примером этого может служить удивительное сходство жилкования крыльев верблюдок и златоглазок, иногда разносимых по разным отрядам. Поскольку характер жилкования, а тем более небольшие его детали, если и влияет на аэродинамику крыла, то не особо значительно (ведь существует огромное количество типов жилкования даже у видов, летающих сходным образом), а также исходя из принципа вероятности, можно сделать вывод, что параллельное развитие практически идентичного жилкования не только в разных отрядах, но и в разных семействах, а то и родах, невозможно, и, следовательно, что верблюдки и златоглазки являются ближайшими родственниками. Три описанных выше принципа можно сформулировать в виде одного афоризма: «Лень — двигатель эволюции».
Интересен также принцип эволюционных тенденций, согласно которому моно- либо полифилетически может возникать не только какой-либо конкретный признак, но и тенденция к его возникновению (эволюционную тенденцию можно, таким образом, рассматривать как самостоятельный биологический признак), причём чем ближе филогенетически находятся таксоны, тем больше сходство в их эволюционных тенденциях. Механизмы такой «эволюции тенденций» остаются неясными (возможно, они кроются в «спящих генах», а может быть, дело в закономерностях между планом строения и дальнейшей эволюцией [Северцев, 90]), но примеры этого многочисленны, на что обратил внимание ещё Л. Берг в гипотезе номогенеза [Грант, 80]. Так, эволюция Lamellicristata (организмов с пластинчатыми кристами митохондрий) наблюдаются тенденции к многоклеточности (животные, растения, грибы, багрянки; за пределами Lamellicristata — только бурые водоросли из Tubulicristata), к выходу на сушу (животные — неоднократно, растения, грибы; за пределами группы — только различные слизевики, предпочитающие, однако, влажные места) [Мирабдуллаев], хотя сами эти признаки (многоклеточность и сухопутность), несомненно, полифилетичны. Подобным образом архозавры имели тенденцию к двуногости (даже крокодилы и гигантские четвероногие динозавры, по некоторым данным, являются четвероногими вторично) и к полёту (птерозавры, птицы) [Наумов, Карташёв,79, т. 2, стр. 13-14].
Принцип холизма заключается в том, что целое есть не сумма составляющих его частей, а другая форма организации материи. Иными словами, элементарные частицы, атомы, молекулы, клетки и живые организмы есть не разные уровни организации, а всего лишь количественно различные, но равные качественно формы (холоны). Исходя из этого принципа, можно сделать вывод, что эволюция неорганического и органического мира основывается на одних и тех же законах, модифицированных для данных объектов [Московский, 99].
Принцип направленности заключается в том, что система живых организмов существует объективно, а эволюция только заполняет имеющиеся в ней вакансии [Московский, 99].
Из принципа направленности вытекает и принцип допустимости, который говорит о том, что эволюционные изменения происходят в некоем допустимом вероятностном пространстве.
К принципам направления относится также принцип сопряжённости, согласно которому эволюционные преимущества имеют не только те организмы, которые побеждают в борьбе за существование сами по себе, но и те, которые наиболее способствуют эволюции экосистемы, биосферы и/или вселенной в целом. Сопряжённость эволюции связана с предопределённостью и объективностью системы живых организмов [Вернадский, 60; Молчан, 98].
1.4. Принципы систематизации
В отличие от ПО, описывающих действие самого эволюционного процесса, и ПДМ, пригодных для применения метода к любому материалу (а не только биологическому), ПС служат для составления удобной и филогенетически верной системы. К принципам систематизации здесь отнесены принципы детерминирования, кладизма, унификации и повышения статуса.
Принцип детерминирования заключается в том, что чем раньше в филогенезе или, соответственно, в онтогенезе закладывается тот или иной признак, тем большее систематическое значение он имеет. Исходя из этого принципа, наиболее важно в классификации животных строение сперматозоидов и яйцеклеток, далее идёт характер дробления, далее — тип гаструляции, способ закладки целомов и т.д. [Адрианов, Малахов, 85; Беклемишев, 79; Захваткин, 49; Иванов, 68; Малахов, Реунов].
С точки зрения принципа кладизма, новая систематическая группа возникает не с достижением предковой группой определённого уровня развития, свойственного этой новой группе, а с появлением предпосылок к возникновению характерной для данной группы организации [Павлинов, 90]. Пользуясь этим принципом, можно избежать типичной для современной систематики проблемы, когда, с одной стороны, трудно разделить ископаемые формы между предковой и потомковой группой, так как одни возможные предки близки к потомкам по одному признаку, а другие — по другому, а с другой — избежать полифилетичности и, в какой-то мере, парафилетичности таксона. Характерный пример: при выделении группы Mammalia сомнительно систематическое положение некоторых тероморф, тогда как при выделении монофилетичной группы Sinapsida эта проблема отпадает.
Принцип унификации вытекает из самой идеи единства живой природы и предполагает, что к любым систематическим группам применимы одни и те же критерии классифицирования, хотя не для всех групп эти критерии найдены. Из этого принципа, в частности, следует необходимость согласования зоологический и ботанической классификации и таксономии (или, если быть точнее, необходимость создания единой биологической систематики и таксономии; каковая, впрочем, в настоящее время как раз и создаётся на основании сравнительного молекулярного анализа), а также потребность в оценке значимости признаков (что можно сделать, основываясь, например, на других ПС, а также с учётом механизмов эволюционных преобразований).
Принцип унификации следует согласовывать с принципом повышения статуса, гласящим, что с повышением уровня организации повышается общая значимость систематических признаков. Так, в основе деления на царства наиболее примитивной группы живых организмов — вирусов — лежит такой глобальный признак как характер наследственного материала и стратегия его реализации [Жданов,66, стр. 49], тогда как у клеточных форм жизни этот признак един для всех (наследственность обусловлена двойной спиралью хромосомной или хромонемной ДНК, причём информация реализуется с помощью т- и иРНК), а деление на царства происходит на другом основании; выделение типов внутри прокариотических царств основывается на особенностях клеточного метаболизма, тогда как, например, внутри царств Zoobionta и Phytobiontaметаболизм более-менее однообразен.
Приведённые выше ПУМ (хотя перечень их, по-видимому, не исчерпывающ) являются не только основными принципами умозрительного подхода к теории эволюции, но и (особенно ПО) важными эволюционными принципами. Основываясь на ПУМ и имеющихся у нас в распоряжении фактах, можно выделить основные эволюционные механизмы и выстроить возможную картину эволюции в живой природе. При этом следует учитывать соответствие различных гипотез как можно большему числу описанных ПУМ.
-
Возникновение жизни
Жизнь можно определить как способ взаимодействия материи, энергии и информации, характеризующийся способностью системы к самовоспроизведению по единой информационной матрице (информационный критерий), обмену веществом, энергией и информацией с окружающей средой (экологический критерий), а также индивидуальному и историческому развитию вследствие случайных или закономерных изменений в информационной структуре (эволюционный критерий). Информационный критерий имеет следствием рождение, умирание и наследственность, экологический — дыхание, питание, выделение, рост и раздражимость, эволюционный — изменчивость, онтогенез и филогенез. На наш взгляд, эти три критерия являются необходимыми и достаточными для понимания сущности жизни, и введение четвёртого, химического критерия (белковая или нуклеопротеиновая природа) только неоправданно сужает поле биологических исследований, не давая потенциальной возможности для изучения небелковых форм жизни(прежде всего инопланетных), если таковые будут обнаружены. Напротив, ликвидация любого из трёх вышеперечисленных критериев ведёт к неоправданному расширению поля исследований и включению в него явно лишних объектов. Так, без информационного критерия живыми существами можно было бы назвать биогеоценозы, планеты, звёзды и т. д., что хотя и может оказаться логичным с мистической точки зрения, но не оправдано биологически; без экологического критерия к живым существам можно было бы отнести большинство информационных объектов (человеческие мысли, компьютерные базы данных и т. п.), что тоже согласуется с некоторыми мистическими представлениями, но не с биологией; наконец, без эволюционного критерия живыми существами являлись бы машины, сконструированные для изготовления себе подобных, что является пока что объектом хотя и фантастики, но вполне научной. Приведённое же выше определение жизни позволяет по-новому взглянуть на проблему происхождения жизни на Земле, до сих пор являющуюся одним из важнейших дискуссионных вопросов теории эволюции.
В настоящее время наибольшей популярностью пользуются гипотезы химического (автохтонного) и космического (панспермия) появления жизни на Земле (именно на Земле, так как космическая гипотеза не снимает вопроса о происхождении жизни вообще) [Васильев, 65; Войнович, 88], причём традиционные биохимические теории [Опарин, 57, 66] сталкиваются с всё большими затруднениями, тогда как космические гипотезы получают всё новые подтверждения. Так, уже доказана возможность переноса сложных органических соединений через космическое пространство на метеорных телах, а также возможности ряда микроорганизмов выдерживать условия открытого космоса в анабиотическом состоянии. Сравнительно недавно получила подтверждение гипотеза, согласно которой жизнь может существовать в недрах практически любой планеты, так как, во-первых, по мнению американского астробиолога Д. Ноэвера и английского генетика С. Джонеса, земные условия можно считать уникальными только в том случае, если рассматривать поверхность, а не недра, во-вторых, в нефти обнаружены такие своеобразные по своей биохимии организмы как археи, а, в-третьих, существует гипотеза, что нефть образует не отдельные скопления в толщах литосферы, а особый слой в строении Земли (и, возможно, других планет), причём, по мнению американских астрономов Дж. Клампета и Т. Голда, нефть отнюдь не является продуктом жизнедеятельности растений, как это предполагалось ранее [Всё живое на Земле произошло из нефти! 99]. В таком случае, жизнь (или, во всяком случае, предбиологические системы типа нуклеиновых кислот) может оказаться почти таким же древним явлением, как и Вселенная.
Как уже было сказано, современные гипотезы химического происхождения жизни на Земле страдает рядом внутренних противоречий, которые свидетельствуют о маловероятности такого пути возникновения живых организмов. Так, можно отметить, что «опаринским» путём сложные органические соединения могли возникнуть только в восстановительной среде, а сохраниться — только в окислительной, что создаёт главный парадокс. Второй парадокс, подобный вопросу «о курице и яйце», заключается в том, что синтез нуклеиновых кислот невозможен без белков, тогда как белки кодируются нуклеиновыми кислотами. Парадоксальным является также, например, построение всех белков только из двадцати аминокислот из более сотни известных, причём из их левоасимметричных форм (вариант решения этой и некоторых других проблем указан в дополнении к данному разделу), и многие другие моменты. Кроме того, вода является сильным растворителем, что тоже говорит не в пользу «гипотезы первичного бульона» (во всяком случае, не в её традиционном представлении).
Не претендуя на научность аргументации, мы можем предложить гипотезу появления жизни на Земле, представляющую собой синтез космической и химической гипотез с теорией последовательных эндосимбиозов, применённой не только к формированию эукариотического типа организации клетки, но и к возникновению прогенота. Так называемые «споры жизни», фигурирующие в гипотезе панспермии (назовём ихвитоспорами), могли представлять собой систему, включающую нуклеиновую кислоту (ДНК либо, вероятнее, РНК) и белковую или липопротеиновую оболочку [Маргелис, 83, стр. 19] и способную при попадании в подходящие условия начать процесс самокопирования, то есть наделённую информационным признаком жизни; таким образом, витоспоры могли напоминать вирион.
К определённому моменту (около 3,5-4 млрд. лет назад) химическая эволюция на Земле привела к возникновению коацерватоподобных образований, способных к обмену веществ и росту, то есть имеющих экологический признак жизни, но неспособных к копированию входящих в их состав молекул [Опарин, 57, 66]. В результате бомбардировки земной поверхности ДНК- и/или РНК-содержащими (первичными «молекулами жизни» могли быть и пептидо-нуклеиновые кислоты, ПНК [Тарасов]) витоспорами различные разновидности последних (или же витоспоры единственного типа, впоследствии эволюционирующие внутри клетки) неоднократно вступали в эндосимбиозы с присутствующими на Земле коацерватами, что позволило совместить информационный и экологический критерии живой материи и привело к появлению третьего компонента — способности к эволюционированию. Вероятно, предбиологическая эволюция органического вещества на Земле и формирование симбиоза витоспор и коацерватов во многом была связана с вращением галактики, в ходе которого периодически менялся уровень космической радиации, влияющей на скорость эволюционного процесса [Шкловский, 73; Назаров, 91, стр. 117]. С вращением же галактики может быть связано и периодическое попадание различных планет в потоки вещества, несущего витоспоры.
Механизм первоначальной симбиотической деятельности коацервата и витоспор неясен, однако очевидно, что ДНК-витоспоры дали начало нуклеоиду прогенота (и через него — геному бактерий, архей, эукариот и их прокариотических эндосимбионтов), а РНК-витоспоры (возможно, разные их формы) — матричным, транспортным и рибосомным РНК клеток (вероятно, первичными были структуры, напоминающие тРНК) и всему рибосомному комплексу, отличающему клеточные формы жизни от неклеточных (вирусов) (процесс симбиогенного возникновения прогенота и эукариот, а также возникновение основных форм жизни — см. рис. 1). Впрочем, не исключена и несколько иная картина: витоспоры могли содержать только один тип нуклеиновой кислоты (одноцепочечная или двухцепочечная ДНК или РНК), а другие типы могли возникнуть уже в результате земной эволюции витоспоры (самостоятельной или, скорее, в комплексе коацерват-витоспора). Эта версия косвенно подтверждается явлением прямой и обратной транскрипции [Вирусология, 89; Жданов, 66], а также существованием сложных поксвирусов — вероятных потомков коацерватов с одним типом нуклеиновой кислоты, — самостоятельная же эволюция витоспор кажется нам маловероятной.
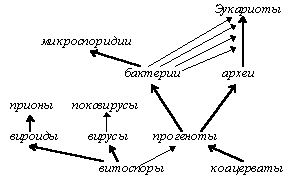
С момента объединения витоспор и коацерватов в единый симбиотический комплекс началось развёртывание на Земле эволюционной программы, подобной космической эволюции химических элементов как заполнению вакансий периодической системы от водорода к тяжёлым элементам. Говоря об «эволюционной программе», мы не имеем в виду, что изначально было определено всё нынешнее многообразие видов (эта гипотеза полностью отвергала бы действие окружающей среды и случайных факторов): мы считаем лишь, что ещё не раскрытые человеком законы природы направляют эволюцию в любых условиях по одним и тем же магистральным путям, подобно тому, как периодическая система отнюдь не находится в противоречии с фактом существования изотопов. Однако точно убедиться в правильности того или иного взгляда на эволюцию в космическом масштабе мы сможем только в том случае, если обнаружим более или менее высокоорганизованную нуклеопротеиновую (или иную) жизнь на других планетах.
Дополнение к разделу 2
Выше говорилось, что синтез белков по матрице нуклеиновых кислот вряд ли мог возникнуть в земных условиях. В последнее время мы заинтересовались этапами ранней эволюции жизни, связанными с такими глобальными явлениями как возникновение разных типов аминокислот (в частности, их левосторонней асимметрии) и нуклеотидов, а также генетического кода. Не вдаваясь в подробности хода наших мыслей, приведём здесь некоторые сделанные нами выводы по этому вопросу, хотя они и требуют ещё дальнейшей доработки (особенно по биохимическому объяснению нижеперечисленных событий).
- Левосторонняя асимметрия аминокислот, во многом создавшая условия для появления белковой жизни, может объясняться единственным способом, не вступающим в противоречие с теорией вероятности: все левосторонние «волшебные аминокислоты», входящие в состав белков, происходят от единственной левоасимметричной «волшебной аминокислоты» простого строения в результате ряда своеобразных «химических мутаций».
- Первичная нуклеиновая кислота состояла только из одного типа нуклеотидных мономеров, кодирующих единственную аминокислоту (по-видимому, это были нуклеотидные остатки цитозина), и имела (в том случае, если первичная нуклеиновая кислота была двухцепочечной) либо синтезировала (если она была одноцепочечной) негативную (не являющуюся матрицей) цепочку комплиментарных нуклеотидных остатков.
- Комплексы нуклеиновой кислоты и аминокислоты напоминали, вероятно, современные транспортные РНК.
- Параллельно идущие «химические мутации» нуклеотидов и аминокислот постепенно привели к возникновению современного их числа.
- Наиболее простая мутация процесса матричного синтеза привела к считыванию информации с негативной нити РНК или ДНК, либо связанной, либо не связанной с позитивной пространственно.
- Пять главных нуклеотидов (а также их аномальныеформы ) сформировались раньше, чем двадцать «волшебных аминокислот).
- На ранних этапах эволюции жизни аминокислоты кодировались двумя, а не тремя нуклеотидами. Третий нуклеотид триплета представлял собой аналог современных генетических «знаков препинания», но не между генами, а между кодонами. В это время белки состояли только из 7-9 типов аминокислот (тогда как максимальное количество аминокислот, которые можно закодировать дуплетным кодом, равно шестнадцати).
Эти и другие выводы можно сделать, основываясь на сопоставлении структуры триплетов современного генетического кода, однако для более полной научной проработки данного вопроса нужно более подробно ознакомиться со структурами и свойствами нуклеиновых кислот и аминокислот. Решение этой проблемы позволит описать возникновение нуклеопротеиновой жизни как таковой, а не только в масштабах земной биосферы.
- Уровни и механизмы эволюции
3.1. Уровни эволюции
В настоящее время принято разделять эволюционный процесс на два уровня — микро- и макроэволюцию; при этом микроэволюцией обычно называется эволюция на уровне популяций, ведущая к видообразованию, а макроэволюцией — эволюция на надвидовом уровне [Грант, 80, 91; Назаров, 91; Черникова, 87]. Однако это деление в значительной степени условно, и прежде всего в связи с тем, что до сих пор не существует точного и всеобъемлющего определения вида (все существующие определения имеют ряд ограничений: например, для видов без полового размножения, видов-двойников, видов с кольцевыми ареалами и т.п. [Грант, 80]; попытка универсального определения видов по установлению мономорфных видоспецифичных участков генома [Назаров, 91, стр. 245] пока что не получила практического распространения), а тем более — крупных таксонов. Поэтому, вероятно, следует разграничить уровни эволюции, основываясь на других принципах, нежели на заведомо условных таксономических, например, на эволюционных механизмах, характерных для того или иного уровня. В пользу такого предположения говорит и гипотеза Мейена о предопределённости эволюции и независимости систематики от количества исследованных групп [Московский, 99].
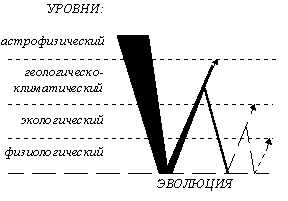
Имеется также и другая концепция подразделения эволюционного процесса: по уровням биологической организации, затрагиваемым данными уровнями преобразования живой материи. Основываясь на таком принципе, историческое развитие жизни подразделяется на онтогенез (изменения на организменном уровне), филогенез, включающий микроэволюцию (изменения на популяционно-видовом уровне) и макроэволюцию (изменения на уровне видов и выше, с момента возникновения генетической изоляции), и сингенез (изменения на биосферно-биогеоценотическом уровне) [Назаров, 91, стр. 11-12]. Процессы масштабом ниже онтогенетического попадают в компетенцию физиологии, биохимии, генетики и т. п., а выше сингенетического — в компетенцию астрономических наук (см. табл. 2). Теория же эволюции должна затрагивать и онто-, и фило-, и сингенез, и даже пограничные области соседствующих с ними в этой системе дисциплин, касающихся, в основном, циклических процессов развития.
Таблица 2. Сфера компетенции общей теории эволюции
| Этапэволюционирования | Сферакомпетенции | Науки |
| космогенез | космические процессы | астрономия |
| сингенез | историческое развитиесообществ и биосферы | эволюционнаяэкология |
| филогенез | историческое развитиеживой природы | теорияэволюции |
| онтогенез | индивидуальное развитиеорганизмов | эмбриология,биология
постнатального онтогенеза |
| жизнедеятельностьорганизмов | физиология,генетика,
биохимия и пр. |
По мотивам, которые будут понятны из дальнейшего изложения, филогенез разделён нами на три уровня: микро-, мезо- и макроэволюция (или, во избежание омонимии, мегаэволюция), причём их границы несколько отличаются от традиционно принятых (см. рис. 2) и не совпадают с границами тех или иных традиционно выделяемых систематических групп: микроэволюция, имеющая в своей основе «дарвиновские», тихогенетические механизмы эволюции [Дарвин, 52], затрагивает внутривидовые таксоны, виды, а в крайнем случае — роды; мезоэволюция, основой которой являются макромутации (прежде всего, связанные с изменением числа хромосом — как кратного, так и некратного), мутации регуляторных генов, приводящие к гетеротопиям и гетерохрониям типа акцелерации, гиперморфоза, прогенеза и неотении [Назаров, 91, 205], и горизонтальный перенос подвижных элементов ДНК [Тихоненко, 80; Гвоздёв, 98; Глазер, 98; Назаров, 91, стр. 41; Хесин, 84], влияют на образование новых родов, семейств, отрядов и групп более высокого порядка (возможно, иногда — до типа включительно); наконец, мегаэволюция, механизмы которой связаны с изменением энергидности и степени самостоятельности субъектов биологических конгломератов, ведёт к образованию типов, царств и более крупных групп и основывается на таких явлениях как колониализация, межвидовая гибридизация, симбиогенез, автономизация клеточных элементов и т. п. (см. табл. 3).
Элементарным процессом микроэволюции являются мутации и рекомбинации, мезоэволюции — генетическая изоляция (как первичная, являющаяся непосредственным следствием мутаций, так и вторичная, вызванная другими типами изоляции), а мегаэволюции — мегаэволюционная детонация, явление, которое будет описано в дальнейшем. Микроэволюция связана с изменением генетической структуры, мезоэволюция — с преобразованием общей схемы организации [Назаров, 91, стр. 14], а мегаэволюция — с изменением энергидности. Помимо этих трёх уровней филогенеза, будут рассматриваться также некоторые вопросы, касающиеся сингенеза в планетарном и космическом масштабе.
Таблица 3. Уровни эволюции
| Уровни | микроэволюция | мезоэволюция | мегаэволюция |
| Таксоны | популяции,сорта, породы,
виды, роды |
роды, семейства,отряды, классы,
типы |
типы, царстваи выше |
| Изменения | медленные,постепенные,
случайные |
мгновенные,случайные | мгновенные,но с постепенным
повышением уровня облигатности |
| Механизмы |
структурных генов
существование
отбор |
(в том числе полиплоидия и анеуплоидия)
регуляторных генов, ведущие к гетеро- хрониям и гетеротопиям
перенос генов |
гибридизация
клеточных структур |
| Направление | ненаправленныйпроцесс | аллогенез | арогенез,катагенез |
| Результаты | разнообразиеродственных
форм |
генетическаяизоляция | новые формыи уровни органи-
зации жизни |
| Ограничения | допустимыепределы
вариаций вида |
летальностьбольшинства
макромутаций? |
биологическаяцелесообразность |
А В

Как можно заметить из таблицы, «мишени» различных уровней эволюции в какой-то степени пересекаются друг с другом. Это связано, однако, не с тем, что на одном и том же уровне действуют разные механизмы, а со значительной условностью современной систематики, в которой аналогичным образом возникшие группы могут иметь разный таксономический статус. Что интересно, эти три уровня эволюции, по-видимому, не пересекаются: вопреки традиционному мнению, мезоэволюция и мегаэволюция отнюдь не складываются из множества микроэволюционных изменений, а протекают совершенно независимо [Четвериков, 26; Назаров, 91, стр. 30]. Именно такой вывод можно сделать из множества фактов, когда тот или иной орган имеет полезные свойства только во всей своей сложности и, следовательно, не мог возникнуть путём постепенного усовершенствования (впрочем, многие подобные факты можно объяснить с точки зрения преадаптации).
3.2. Особенности микроэволюции
Mеханизмы, отнесённые в данной работе к механизмам микро- и мезоэволюции, хорошо известны в теории эволюции. Однако если механизмы микроэволюции (микромутации, рекомбинации, борьба за существование, естественный отбор, поток генов и дрейф генов) и по традиционным учениям относились именно к микроэволюции и видообразованию [Грант, 80, 91], то механизмы, отнесённые здесь к мезоэволюции, были разделены между процессом видообразования и макроэволюции.
Поскольку учение о микроэволюции не слишком сильно изменилось со времён Дарвина, а её механизмы хорошо изучены теоретически и практически (например, путём сравнения родственных видов-эндемиков или видов с кольцевыми ареалами наподобие Larus argentatus-fuscus), здесь микроэволюция рассматриваться не будет. Здесь же следует только дать определение микроэволюции как процесса постепенного ненаправленного преобразования видов. При этом следует отграничивать микромутации как механизм, формирующий микроэволюционные изменения (новые аллели), и рекомбинации как механизм, образующий новые их комбинации, от других общеизвестных механизмов, регулирующих лишь соотношение аллелей в популяции. Эти механизмы можно объединить под общим названием движущие силы микроэволюции.
Следует отметить, что, в отличие от эволюции более крупных уровней, микроэволюция является обратимым процессом. Действительно, точковые микромутации, не ведущие к резкому изменению выживаемости и размножаемости, возникают случайно, и так же случайно могут возникнуть мутации, им противоположные (прямые и обратные транслокации и инверсии, дупликации-делеции и т. п.); борьба за существование и естественный отбор, хотя и не случайны по своей природе, ведут только к относительной приспособленности к данным условиям, поэтому при случайном изменении условий сначала в прямом, а затем в обратном направлении тоже будут возвращать популяцию к исходным формам; поток генов и дрейф генов — случайные явления, к тому же, поток генов ведёт к изменению соотношения аллелей в пределах популяции, а не вида. Иногда говорят даже, что микроуровень эволюции вообще является не собственно эволюцией, как мезо- и мегауровень, а адаптивной внутривидовой дифференцировкой [Назаров, 91, стр. 41].
Впрочем, микроэволюция тоже может вести к изоляции вида, однако изначально это будет обычно не генетическая, а какая-либо другая форма изоляции (географическая, экологическая и т. п.). Подобным способом, например, возникли уже упомянутые L. argentatus и L. fuscus [Назаров, 91, стр. 29-30]. Таким образом, полностью отрицать видообразовательное (и макроэволюционное в традиционном понимании) значение микроэволюционных процессов мы, безусловно, не можем. Тем не менее, нам представляется более распространённым сальтационное (мезоэволюционное) видообразование с первичностью генетической изоляции.
3.3. Понятие и механизмы мезоэволюции
Мезоэволюцию можно охарактеризовать как эволюционный процесс, ведущий ко внезапной (в течение одного поколения) генетической изоляции без предварительного отбора. Как уже было сказано, основными механизмами мезоэволюции являются макромутации, мутации регуляторных генов и горизонтальный перенос генов.
Одним из наиболее часто встречаемых (в основном, среди растений) способов возникновения систематических групп мезоэволюционного уровня является полиплоидия, ведущая к мгновенному (без предварительного накопления каких-либо «полезных признаков», что ещё раз свидетельствует об отграниченности мезоэволюционного процесса от микроэволюционного) возникновению репродуктивной изоляции и, соответственно, к образованию нового вида [Эволюция генома, 86] (возможно, более точно следует считать это событие возникновением нового рода, так как здесь будет отсутствовать даже возможность появления стерильных гибридов с предковой формой, имеющее место при скрещивании видов одного рода, как в случае с родом Equus). Впрочем, полиплоидизацию можно отнести и к мегаэволюции, которая по определению связана с изменением энергидности.
Помимо полиплоидии, изменение числа хромосом, ведущее к видообразованию, может происходить и другими путями. Так, может произойти удвоение не всего набора хромосом, а только части хромосом (анеуплоидия); кроме того, может иметь место утрата одной или нескольких хромосом, разделение одной хромосомы на несколько или слияние нескольких в одну в результате неправильно протёкшего процесса митоза. В большинстве случаев, разумеется, такие изменения в генотипе являются летальными или ведущими к стерильности, однако несомненно, что иногда подобные преобразования могут привести и к возникновению новых видов. Так, по-видимому, возникновение разных видов приматов (в том числе человека), имеющих разное число хромосом, но поразительное сходство в синтезируемых белках, связано именно с такими изменениями числа хромосом [Назаров, 91, стр. 40].
Фактическая гомологичность хромосом между собой и с витоспорами свидетельствует о том, что и поли-, и анеуплоидию, и объединение или разделение хромосом можно отнести и к мегаэволюционным механизмам, тогда как не особенно крупные по масштабам воздействия на генотип мутации регуляторных генов могут привести к явно мезоэволюционным преобразованиям. Наиболее существенно в этом плане явлениегетерохронии (в частности, неотения), происходящее в результате мутаций генов, регулирующих сроки полового созревания. Хотя генетическая перестройка при неотении небольшая, она позволяет «сбросить груз специализации» и, таким образом, снимает действие принципа тупиковости специализации, направляя эволюцию в горизонтальном (неотеническом) и вертикальном направлении (см. рис. 3); наиболее ярким примером этого процесса мы можем считать эволюцию животных. Кроме того, неотения приводит к мгновенной репродуктивной изоляции, хотя в некоторых случаях (классический пример — амблистома и аксолотль) неотения может быть факультативной. Гетерохронии типа прогенеза и неотении ведут к педоморфозу, а гетерохронии типа акцелерации и гиперморфоза — к рекапитуляциям [Назаров, 91, стр. 205]. Гетеротопии, в отличие от гетерохроний, ведут к изменению не времени, а места закладки органов, либо к изменению их пространственной ориентации. Как и гетерохронии, гетеротопические изменения сыграли существенную роль в эволюции высших таксонов животных.
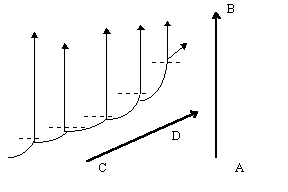
Интересным и своеобразным механизмом мезоэволюции могут являться так называемые онтомутации — мутации материнской иРНК, контролирующей дробление яйцеклетки и начальные стадии онтогенеза. Поскольку онтомутации возникают у зародыша на начальных стадиях развития, они могут не вести к тяжёлым последствиям для организма и одновременно способны в течение одного поколения резко изменить план его строения. Кроме того, гипотеза об онтомутациях снимает вопрос о происхождении нового таксона от единственного «обнадёживающего урода», так как схожие мутации иРНК, в отличие от мутаций в генотипе, могли бы возникать у значительной части популяции под влиянием одного и того же фактора [Назаров, 91, стр. 151-157].
Все макромутационные изменения могут происходить на генах, возникших в результате дупликации, в том числе множественной, что не только снижает вероятность их летальности, но и ведёт к общему увеличению содержания ДНК, а также к возникновению принципиально новых генетических локусов, и может способствовать прогрессивной эволюции [Назаров, 91, стр. 236]. Тот же результат дают, помимо дупликации генов, и другие способы увеличения количества генетического материала, такие как уже упомянутые поли- и анеуплоидия.
В качестве механизма мезоэволюции можно указать также явление горизонтального переноса подвижных участков ДНК с помощью вирусов (вирусная трансдукция) и другими способами (с помощью эпигенов, транспозонов и мобильных генетических элементов [Назаров, 91, стр. 41; Хесин, 84]). Его особенностью является обмен генетической информацией между любыми видами живых организмов без каких-либо ограничений степенью родства [Тихоненко, 80; Гвоздёв, 98; Глазер, 98; Жданов, 66, стр. 42].
Важным для мезоэволюции явлением (не механизмом мезоэволюции, а именно явлением, с ней связанным) можно назвать преадаптации, которые могут возникать либо как второстепенные признаки, сопутствующие полезным преобразованиям, либо в результате мутаций, не ведущих к изменению выживаемости в момент их возникновения, но приводящих к улучшению приспособленности в дальнейшем, либо в результате высвобождения органа от ранее выполняемой функции [Назаров, 91, стр. 74]. Примеры таких преадаптаций можно найти опять-таки среди приматов. Так, в результате обеднения ландшафтов перейдя от брахиации к наземному прямохождению, предки человека сохранили вертикальное положение тела, что позволило им легче приспособиться к условиям открытого пространства. Хватательная конечность, сформировавшаяся у приматов для облегчения передвижения по деревьям, при переходе к наземному образу жизни освободилась для выполнения других нагрузок. Таким образом, преадаптациями можно назвать всякие признаки, не возникающие в результате изменения внешних условий, а помогающие к ним приспособиться, приобретая новые функции [Георгиевский, 74], что опять-таки подчёркивает, что микроэволюционные законы мало уместны для описания мезоэволюции.
3.4. Понятие и механизмы мегаэволюции
Термин «мегаэволюция» был предложен Дж. Симпсоном для обозначения наиболее крупномасштабных преобразований, связанных с возникновением высших таксонов, начиная с семейства или отряда, но позднее Симпсон отказался от него [Назаров, 91, стр. 14; Симпсон, 48, стр. 156], поэтому использование его в несколько других рамках не приведёт к двусмысленностям.
Рассматривая симбиогенную и колониальную теории происхождения эукариот, переход от факультативности к облигатности в симбиозе гриба и водоросли (лишайники), колониальный путь перехода от одноклеточности к многоклеточности у животных и растений, гипотезу автономизации клеточных и генных структур в эволюции вирусов и другие аналогичные примеры, можно прийти к выводу, что мегаэволюционные изменения связаны с изменением степени самостоятельности биологических объектов и, как следствие, с возникновением новых форм и уровней организации живого. При этом очевидно, что в большинстве процессов, являющихся мегаэволюционными механизмами (автономизация, симбиогенез, колониализация), происходит постепенная облигатизация взаимоотношений, сложившихся между исходными бионтами в результате единственного и мгновенного события (например, клеточного каннибализма в случае формирования полового процесса, пожирания предэукариотической клеткой прокариотических эндосимбионтов, отделения участков ДНК при возникновении вирусов и т. п.). Это событие названо здесь мегаэволюционной детонацией и представляет собой элементарный процесс мегаэволюции, подобно тому, как генетическая изоляция (и, как следствие, видообразование) является элементарным процессом мезоэволюции, а точковая мутация — микроэволюции.
Во многих случаях мегаэволюционных преобразований (утрата самостоятельности пластидами или симбионтами-компонентами лишайников) мы можем проследить постадийность этого процесса, в других же (переход к многоклеточности) такое отслеживание будет носить характер умозрительного. Все эти процессы объединены тем, что в них происходит либо разделение одного бионта n-порядка на несколько бионтов (n-1)-порядка (автономизация вирусов), либо, напротив, слияние далёких друг от друга (симбиогенез и межвидовая гибридизация) или однородных (становление полового процесса, колониализация) бионтов n-порядка в бионт (n+1)-порядка; иными словами, в процессе мегаэволюции бионт приобретает уровень (n± 1) (таким образом возникали, по-видимому, разные подцарства растений и животных, многие царства и систематические группы более высоких рангов). Мегаэволюцию, таким образом, можно охарактеризовать как процесс изменения уровня организации бионтов (или степени их энергидности, что практически равнозначно). Если микроэволюция ограничена допустимыми пределами изменения вида, а мезоэволюция — летальностью большинства преобразований, могущих привести к мезоэволюционным изменениям, то мегаэволюция ограничена, по-видимому, только биологической целесообразностью.
Из возможных механизмов мегаэволюции отдельно стоит сказать о межвидовой гибридизации, как естественной, так и искусственной. К мегаэволюционным механизмам это явление причислено потому, что в нём происходит слияние двух бионтов в один, но этот механизм имеет и существенное отличие от других: здесь отсутствует постепенная облигатизация, свойственная, например, симбиогенезу, колониализации или автономизации. Иными словами, межвидовая гибридизация (а также процессы, связанные с изменением числа хромосом и отнесённые в данной работе к мезоэволюционным механизмам) целиком представляет собой акт мегаэволюционной детонации. В настоящее время доказана гибридная природа многих групп животных и растений [Назаров, 91, стр. 4; Стеббинс, Айала, 85], что подчёркивает огромное значение этого механизма.
3.5. Особенности сингенеза
Сингенезом называется процесс исторического развития экосистем и всей биосферы. К ведению учения о сингенезе относятся, в первую очередь, периодические смены флор и фаун в процессе исторического развития жизни на Земле. Не вдаваясь в геологические, астрономические и другие подробности, отметим здесь только некоторые закономерности этого процесса. Самым существенным тут является зависимость менее масштабных явлений от более масштабных и явление эволюционного отражения. Исходя из этого тезиса, можно сказать, что три уровня биогенетической эволюции живой природы находятся в зависимости от эволюции природных сообществ, эволюция сообществ — от климатических условий, климат — от глобальных теллурических явлений, а те, в свою очередь, находятся под влиянием космических факторов циклического и ациклического характера. Действительно, модели влияния «космос — абиотические факторы среды — биологические процессы» можно проследить на всех уровнях процессов, связанных с периодическими явлениями космического уровня: суточные изменения, связанные с вращением Земли вокруг своей оси, приливно-отливные процессы, вызываемые лунными циклами, сезонные изменения, основанные на годовом цикле Земли, гелиобиологические явления, вызываемые периодами повышения и понижения солнечной активности и связанные, по-видимому, с 11,8-летним юпитерианским годом, и т. п. Причём очевидно, что чем выше уровень космического цикла, тем больше его влияние на абиотические, а следовательно, и на биотические факторы Земли. Поэтому есть все основания полагать, что наиболее глобальные изменения биосферы связаны с наиболее продолжительными космическими циклами, такими как «предварение равноденствий» (ок. 25 тыс. лет), периоды вертикальных колебаний солнечной системы около галактической плоскости («галактические сутки», продолжительность около 30 млн. лет) и галактический год (вероятно, ок. 130-180 млн. лет) [Назаров, 91, стр. 128; Красилов, 86]. Также в той или иной степени влиять на эволюцию могут периодические явления, связанные с изменением магнитного статуса Земли [Николаев, 91].
Имеет место и обратный процесс, когда меньший по масштабности уровень влияет на больший. Поскольку при каждом переходе на другой уровень масштабность процесса убывает, а обратный процесс возникает в ответ на прямое воздействие, это явление можно назвать эволюционным отражением. В ходе прямого процесса часть энергии, возникающей в ходе эволюции космических объектов, воздействует на геологическо-климатическую эволюцию Земли, та, в свою очередь, влияет на сингенетические процессы, а изменения экосистем вызывает изменение на физиологическом и молекулярно-генетическом уровне, которые и ведут к эволюционным изменениям. Но эволюционные изменения на генетическом уровне, в свою очередь, ведут к изменениям морфологии и экологии, что не может не сказаться на состоянии экосистем, а через них — и биосферы в целом (см. рис. 4); затем происходят дальнейшие затухающие колебания. Из логики подобного построения следует, что это, в свою очередь, влияет и на астрофизические процессы, однако постепенная потеря энергии от уровня к уровню имеет следствием то, что это влияние заметить весьма проблематично. Тем не менее, антропическое воздействие на Землю может сказаться и на космических явлениях, хотя бы в масштабе Солнечной системы.
Сопоставление палеонтологических и астрономических данных позволило проследить имеющую место корреляцию между периодом вертикальных колебаний солнечной системы (галактическими сутками) и пиками наибольшего вымирания в различных группах живых организмов (30+4 млн. лет). Что интересно, в палеозое периодичность вымираний была около 34 млн. лет, а в мезозое и кайнозое уменьшилась до 26 млн. лет[Назаров, 91, стр. 130-131], что может свидетельствовать о постепенном сокращении продолжительности «галактических суток» и, возможно, об относительной молодости Галактики, так как после достижения «среднего возраста» колебания, вероятно, должны затухать. На основании этого, а также исходя из того, что скорость мезоэволюции, по-видимому, возрастает, можно сделать вывод (чисто гипотетический), что скорость мезоэволюции каким-либо образом коррелирует с вертикальными колебаниями Солнечной системы.
Возможно, галактический год, как и год земной, характеризуется своеобразными «галактическими сезонами», которым свойственен больший или меньший уровень радиации и/или какие-либо другие черты, и от этого также зависит эволюционный процесс и другие явления, происходящие на Земле (см. рис. 5). Вероятно, с инерционным замедлением скорости вращения галактики может быть связано постепенное замедление мегаэволюционного процесса. Исходя из этих и подобных закономерностей, можно сказать, что сингенетический процесс, по-видимому, цикличен или, если быть точнее, спирален, то есть последующий виток повторяет предыдущий на более высоком уровне.
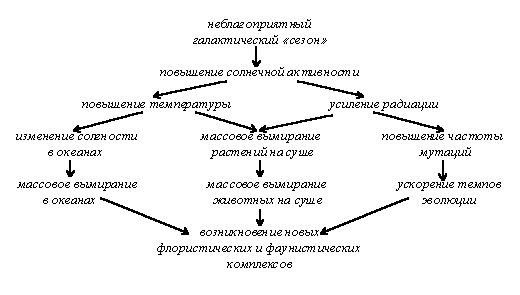
Непериодические явления космического уровня, такие как взрывы сверхновых, ведущие к повышению уровня радиации во много раз [Назаров, 91, стр. 116-118; Красовский, Шкловский, 57, стр. 198], столкновение с Землёй астероидов или комет, непосредственно влияющее на биосферу[Назаров, 91, стр. 121-122], и др., тоже, несомненно, внесли свой вклад в катастрофические изменения природных сообществ Земли. Несомненным образом влияя на эволюцию сообществ, катастрофы (ни экстрателлурические, ни теллурические, — хотя последние, вероятно, тоже связаны с экстрателлурическими факторами), по-видимому, не способствуют напрямую макро- (мезо- и мега-) эволюции, но позволяют вновь возникающим формам с большей вероятностью выжить и занять освободившиеся в результате катастрофы экологические ниши. Тем не менее, процессы, связанные с повышением уровня радиации, даже не особо крупными, могут и непосредственно влиять на эволюцию, повышая скорость вымирания отдельных таксонов и увеличивая частоту мутаций, в том числе и макроэволюционного масштаба. Как и для других уровней эволюционного процесса, для сингенеза характерно явление преадаптации: прогрессивные группы возникают в эпохи господства более примитивных, но более приспособленных к данным условиям групп. При изменении условий и вымирании значительной части представителей господствующей группы на первый план выходит эволюционно более продвинутая группа.
Можно отметить ещё некоторые особенности сингенеза. Так, несоответствие флористических и фаунистических эр и периодов (флористические изменения обычно предшествуют фаунистическим), а также меньшие масштабы вымирания среди растений, чем среди животных, говорят о том, что растения первыми реагировали на космические изменения, но количество способствующих вымиранию факторов для них было меньше, тогда как на животных влияли также изменения во флоре.
3.6. Эволюция процесса размножения
Очевидно, что становление того или иного конкретного способа полового размножения (место редукции, соотношение мужских и женских гамет и т.д.) в различных группах происходило самостоятельно. Однако обычно считают, что, во-первых, разделение на два пола если и полифилетично, то происходило относительно немного раз (например, при формировании царств, о чём свидетельствует, в частности, своеобразие полового механизма у инфузорий [Беклемишев, 79] и многих настоящих грибов [Гарибова и др., 75, стр. 82; Жизнь растений; Курс низших растений, 81; Лемеза, Шуханов, 94, стр. 138]), а во-вторых, что эволюционируют, в основном, особенности полового процесса, а сама его сущность — слияние мужской и женской гаметы — остаётся неизменной. Тем не менее, ряд фактов (и, прежде всего, различия в способах определения полов даже в пределах одного типа [Айала, Кайгер, 87; Генетика, 91; Медведев, 76]) свидетельствует, что разделение на два пола происходило самостоятельно не только в разных типах одного царства, но и в разных классах (среди позвоночных), а то и отрядах (например, среди насекомых), а следовательно, что эволюция крупных таксонов в значительной степени протекает безо всякой зависимости от полового размножения.
Одним из самых любопытных примеров этого плана является отличие генетических характеристик полов в двух ветвях позвоночных — диапсиды и синапсиды (не говоря о частых случаях перехода одного пола в другой у низших позвоночных — рыб и амфибий): так, у синапсид (млекопитающих) гетерозиготны самцы, а у диапсид (птиц, архозавров, лепидозавров) — самки [Айала, Кайгер, 87; Генетика, 91; Медведев, 76], что свидетельствует о том, что разделение на полы происходило независимо друг от друга, по крайней мере, в диапсидной и синапсидной ветвях позвоночных (этот факт является косвенным свидетельством происхождения синапсид и диапсид от разных групп кистепёрых рыб). Вообще, если рассматривать группы с различными способами определения пола, можно прийти к выводу, что, допустим, в группе, выделяемой в настоящее время как класс или надкласс Insecta, возникновение полов происходило независимо, по крайней мере, четырежды: у перепончатокрылых (определение пола по признаку гапло- или диплоидии), чешуйчатокрылых (гомозиготность у самцов), двукрылых (по крайней мере, у дрозофилы со сложной системой доминантности женских либо мужских признаков) и других насекомых (гомозиготность у самок) [Генетика, 91; Медведев, 76], — а скорее всего, гораздо больше раз.
По-видимому, группы с гомозиготностью самцов или самок возникали в результате сложных мутаций половой хромосомы, тогда как группы с гаплодиплоидным типом определения пола или с системой, подобной системе полов у дрозофилы — в результате анеуплоидии (все эти процессы носят мезоэволюционный характер). Однако существуют группы, где формирование полов имеет основу не генетическую, а, например, экологическую [Генетика, 91; Медведев, 76].
Традиционно принято считать, что половое размножение стимулирует эволюцию, так как лишь у организмов с половым размножением действуют такие общеизвестные механизмы эволюции как дрейф генов и поток генов, а также рекомбинантная изменчивость и кроссинговер[Грант, 80]. Однако можно заметить, что эти явления играют роль только на микроэволюционном уровне. Кроме того, очевидно, что, если разделение на полы возникает, по крайней мере, при эволюции внутри типа, то самые крупные эволюционные изменения происходят без участия полового процесса, причём именно при изучении эволюции классов, типов и выше мы сталкиваемся с наибольшими «белыми пятнами эволюции» и наиболее внезапными преобразованиями [Самовар, 91]. Из этого можно сделать следующий вывод: половой процесс отнюдь не стимулирует эволюцию, а напротив, прекращает мега- и мезоэволюционные процессы и подключает микроэволюционные.
Следует рассмотреть наиболее вероятную эволюционную цепочку при становлении полового размножения (см. рис. 6). Разумеется, наиболее примитивной формой размножения следует назвать бесполое, при котором каждая клетка способна дать полноценную особь (вегетативное размножение). Такую форму размножения мы можем наблюдать у одноклеточных и многих колониальных организмов, а из многоклеточных — у губок, кишечнополостных, многих растений, грибов. Следующая ступень эволюции размножения — размножение с дифференцированными группами клеток (клубнями, «усами» и др.) или отдельными клетками бесполого размножения (спорами, или агаметами), которое можно обнаружить в той или иной форме у всех высших растений, а также у фораминифер [Беклемишев, 79, стр. 25]. Агаметы, возможно, выделились из плотных групп однородных клеток вегетативного размножения, большинство из которых впоследствии утратило функцию размножения и стало выполнять функции спорангиев. В пользу происхождения гамет от генеративных агамет свидетельствуют также такие явления как андрогенез (напр., у кукурузы, наездников) и гиногенез (напр., при скрещивании разных видов карасей), а также возможность вызывания у многих видов искусственного партеногенеза путём стимуляции яйцеклетки. Эти факты говорят об изначальной равноценности обоих типов гамет и о первоначальной факультативности оплодотворения.

Из бесполого размножения с помощью спор напрямую можно вывести первичный партеногенез. Первичность партеногенеза во многих (если не во всех) зарождающихся крупных таксонах решает (вместе с гипотезой онтомутаций) ранее непреодолимую проблему «многообещающих уродов» (макромутантов, у которых возникают крупные фенотипические изменения, не ведущие к летальности и впоследствии, как предполагалось, становящиеся преадаптациями): как им удаётся найти брачного партнёра со схожим генотипом и при этом избежать близкородственного скрещивания в дальнейшем [Назаров, 91, стр. 143], — так как при партеногенетическом размножении такой проблемы просто не существует. Кроме того, партеногенез, хотя и происходит с помощью гамет, можно рассматривать как видоизменённое споровое, или агаметическое, размножение. Косвенно о преемственности бесполого соматического, бесполого агаметического (спорового) и полового размножения свидетельствует также, например, явление соматического эмбриогенеза у губок.
На каком-либо этапе у различных групп организмов, имеющих гаплоидные агаметы, происходило слияние агамет, приобретающих, таким образом, функции гамет. При этом гаметы сперва были одинаковыми морфологически и физиологически (изогамия, а в случае одноклеточных организмов сперва хологамия), затем приобретали физиологические отличия, при которых сливаться могли только гаметы противоположных «знаков», затем одна из гамет становилась больше другой за счёт накопления питательных веществ, и, наконец, возникала классическая оогамия [Петров, 74]. Не исключено, что возникновение полового процесса было связано с адаптацией к клеточному каннибализму, что напоминает мегаэволюционные механизмы (возникновение диплоидной формы жизни из двух гаплоидных). В таком случае, акт каннибализма соотносим с мегаэволюционной детонацией, а после неё следует процесс постепенной облигатизации оплодотворения. Оогамия, судя по всему, возникла в результате мезоэволюции, тогда как гетерогаметизация — процесс явно микроэволюционный.
Разумеется, обнаружить полную цепочку эволюции процесса размножения в развитии какого-либо организма практически невозможно, так как отдельные стадии зачастую выпадают. К тому же, у огромного количества групп имеет место сохранение примитивных форм размножения при наличии прогрессивных. Так, например, кишечнополостные, губки, высшие растения, приобретя способность к половому размножению, весьма часто «пользуются» и вегетативным размножением, многие растения чередуют стадии с половым и споровым размножением, а у такой прогрессивной группы как млекопитающие бесполое размножение сохранилось в видоизменённой форме, в виде деления зиготы с возникновением однояйцевых близнецов (у броненосцев, например, эта форма размножения достаточно распространена).
3.7. Критика концепции искусственного отбора
Отдельно следует сказать о часто рассматриваемом разграничении эволюционного процесса на эволюцию, связанную с естественным отбором (эволюцию биологических видов), и эволюцию, связанную с искусственным отбором (эволюцию сортов и пород). Согласно таким концепциям, человек противопоставляется всей остальной природе (следствие антропоцентрического мировоззрения), тогда как в действительности человек выступает в роли внешнего (по отношению к популяции, в которой производится отбор) фактора, наряду с другими факторами природы. Противопоставление антропогенного (антропического) фактора биотическому в корне неверно с точки зрения естественного классифицирования: хотя он и весьма значителен в планетарном масштабе, но, подходя с качественной, а не с количественной позиции, неотличим от, например, воздействия муравьёв на популяции тлей или от мелиоративной деятельности бобров.
С точки зрения сторонников концепции искусственного отбора, сорта растений и породы животных не подвержены борьбе за существование. Однако в действительности можно сказать, что для них борьба за существование просто проявляется в других формах, нежели для их «диких» родственников, — а именно, в соответствии определённых признаков запросам биологического вида животных Homo sapiens, одного из многочисленных компонентов живой природы.
Таким образом, искусственный отбор представляет собой всего лишь форму естественного, поскольку антропический фактор отличен от биотического только в масштабах и формах воздействия на природу, но не в качественных признаках: и тот, и другой представляют собой изменение окружающего мира живыми организмами (включая человека) с целью приспособления к условиям среды и с помощью своей активной деятельности. Тем более, разделение естественного и искусственного отбора теряет свою актуальность, если признать, что отбор лежит только в основе микроэволюции, так как в этом случае исчезает такое важное отличие как разные результаты действия двух видов отбора.
3.8. Взаимовлияние эволюционных механизмов
Хотя мега-, мезо- и микроэволюционные механизмы действуют самостоятельно, они взаимно влияют друг на друга. Так, на определённом этапе к мегаэволюционным механизмам подключаются мезоэволюционные, а к мезоэволюционным — микроэволюционные. Наиболее важным здесь является уже упоминаемое ранее явление неотении, поскольку, как отмечалось ранее, высокая специализация затрудняет эволюцию, а явление неотении позволяет уменьшить специализацию и, таким образом, усилить действие микроэволюции. Неотения, возникшая у партеногенетически размножающихся организмов (педогенез), является, по всей видимости, базовой на момент возникновения полового процесса [Захваткин, 49]. Кроме того, механизмы всех уровней эволюции оказывают влияние на виды одновременно, хотя и каждый по-своему, и, тем не менее, достаточно просто путём логических умозаключений прийти к выводу, каким путём произошло то или иное преобразование. От этого зависит и систематическая ценность различных признаков.
Можно отметить следующие типы взаимовлияния различных уровней эволюционного процесса. Во-первых, мега- и мезоэволюционные преобразования подвергаются впоследствии микроэволюционному отбору. Во-вторых, завершение действия мегаэволюционных механизмов приводит к подключению мезоэволюционных, а завершение действия мезоэволюционных — к подключению микроэволюционных. В-третьих, как уже отмечалось, некоторые мезоэволюционные преобразования сами по себе могут способствовать усилению действия микроэволюции. В-четвёртых, одно и то же генетическое изменение может привести к изменениям на разных уровнях. В-пятых, мегаэволюционные преобразования (как, например, приобретение предэукариотами бактериальных эндосимбионтов) могут являться преадаптациями для мезоэволюционных преобразований (в приведённом примере — связанных со становлением митоза и других). Наконец, могут быть и иные способы взаимовлияния.
Можно заметить и некоторую постепенность в переходе от микроэволюционных механизмов к мегаэволюционным через мезоэволюционные. С одной стороны, трудно провести чёткую границу между микро- и макромутациями, различие между которыми является одним из разграничивающих моментов между микро- и мезоэволюцией (в ряде случаев это различие количественное, а не качественное), а даже равнозначные микромутации структурных и регуляторных генов ведут к совершенно разным последствиям. Далее, горизонтальный перенос генов, ведя, как и мутации, к моментальным перестройкам в геноме, уже имеет некоторые мегаэволюционные черты, так как связан с информационным взаимодействием разных (в том числе далёких друг от друга) видов. Гибридизация, представляя собой объединение двух бионтов в один, не характеризуется постепенной облигатизацией взаимоотношений. Наконец, симбиогенез в полной мере обладает мегаэволюционными чертами. Аналогичные сопоставления можно сделать и по другим механизмам (колониализация, автономизация и т. д.). Это говорит, однако, не о единстве и взаимной сводимости разных уровней эволюционного процесса, а напротив, о ещё большей его дробности, что даёт основание подразделять эволюцию на ещё большее число уровней, не связанных друг с другом.
Выявление того, с помощью какого механизма возникала та или иная группа живых организмов, позволит унифицировать систематику, так как этот признак является качественным, а не количественным (то есть показывает реальную природу данной группы, а не степень её отличия от ближайших к ней родственных групп или от предковых форм), и, к тому же, свойственен всем без исключения живым организмам. Возможно, использование такого подхода поможет созданию универсальной системы живых организмов наподобие периодической системы химических элементов, созданной Менделеевым [Московский, 99].
Заключение
В данной работе нами были рассмотрены некоторые аспекты применения умозрительного метода к таким общебиологическим дисциплинам, как эволюционная систематика и теория эволюции. Основным нашим вкладом в проблематику является формулирование принципов умозрительного метода в эволюционной науке, разделение макроэволюционного процесса на мезо- и мегаэволюцию, систематизация с учётом этого разделения основных механизмов надвидовой эволюции, введение понятий о мегаэволюционной детонации, эволюционном зеркале в сингенетических процессах и трёх необходимых и достаточных критериях жизни, формулировка космическо-теллурической (расширенной симбиогенной) гипотезы возникновения жизни на Земле, а также некоторые другие, более частные моменты.
Данная работа носит обзорный характер и предназначена, прежде всего, не для специалистов в области биологии, а для широкого круга читателей, интересующихся проблемами эволюции. Первая редакция этой работы создавалась в 1999-2000 году и, соответственно, практически не учитывает многих новшеств в теоретической и практической биологии, появившихся за последнее десятилетие, однако мы посчитали нецелесообразным внесение в неё существенных новшеств, лишь удалив из неё моменты, заведомо не согласующиеся с произошедшими изменениями в науке и нашем мировосприятии. С некоторыми другими сторонами нашего отношения к проблемам эволюции читатель может познакомиться также в наших статьях «О судьбе эволюционной систематики» («Апокриф», вып. 32, apokrif93.a-z-o-t.com/apokrif.php?c=r_bc16t02_12) и «Преадаптация против “нечленимой сложности”» («Апокриф», вып. 42, apokrif93.a-z-o-t.com/apokrif.php?c=r_bi020032_02).
Библиография
- Адрианов А., Малахов В. Сравнительно-морфологический анализ организации головохоботных червей, филогения и система типаCephalorhyncha // Зоологический журнал. — Т. 74, вып. 4-8.
- Айала Ф., Кайгер Дж. Современная генетика — в 3-х т. — М.: Мир, 1987.
- Беклемишев К. Зоология беспозвоночных. — М.: МГУ, 1979.
- Берг Л. Труды по теории эволюции. — Л.: Наука, 1977.
- Богомолов С. Сокращение раннего эмбриогенеза и дивергентная эволюция у некоторых примитивных многоклеточных животных // Сб. «Темп индивидуального развития животных и его изменение в ходе эволюции». — М.: 1968.
- Васильев И. Живое из неживого. — М.: Знание, 1965.
- Вернадский В. Биосфера. — М.: Наука, 1967.
- Вирусология, в 3-х т. / ред. Филдс Б., Найн Д. — М.: Мир, 1989.
- Войнович Г. Возникновение и развитие жизни на Земле. — М.: Наука, 1988.
- Всё живое на Земле произошло из нефти! // Комсомольская правда. — 1999. — 11 марта.
- Гангус А. Эволюция для всех, или Путь кентавра. — М.: Гелиос, 2001.
- Гарибова Л., Горбунова Н., Сидорова И., Сизова И. Низшие растения. — М.: МГУ, 1975.
- Гвоздёв В. Подвижная ДНК эукариот. // Соросовский образовательный журнал. — 1998, №8. — Стр. 8-21.
- Генетика. — М.: 1991.
- Георгиевский А. Проблема преадаптации. — Л.: Наука, 1974.
- Глазер В. Гомологическая генетическая рекомбинация. // Соросовский образовательный журнал. — 1998, №7. — Стр. 13-21.
- Глазер В. Генетическая рекомбинация без гомологии: процессы, ведущие к перестройке в геноме. // Соросовский образовательный журнал. — 1998, №7. — Стр. 22-29.
- Грант В. Эволюционный процесс. — М.: Мир,1991.
- Грант В. Эволюция организмов. — М.: Мир, 1980.
- Догель В. Зоология беспозвоночных. — М.: 1980.
- Дарвин Ч. Происхождение видов. — М.: ГИСХЛ, 1952.
- Жданов В. Эволюция вирусов. — М.: Медицина, 1966.
- Жизнь — как она возникла? — New York: WBATS, 1985.
- Жизнь растений — в 6-и т. — М.: Просвещение.
- Захваткин А. Сравнительная эмбриология низших беспозвоночных. — М.: Современная наука, 1949.
- Зусман И. Вопросы эволюции эмбриогенеза животных. — М.: Знание, 1973.
- Иванов А. Происхождение многоклеточных животных. — Л.: Наука, 1968.
- Кейлоу П. Принципы эволюции. — М.: Мир, 1986.
- Красилов В. Нерешённые проблемы теории эволюции. — Владивосток: 1986.
- Красовский В., Шкловский И. Возможное влияние вспышек сверхновых на эволюцию жизни на Земле. // ДАН СССР. — 1957, т. 116, №2. — Стр. 197-199.
- Курс низших растений. — М.: ВШ, 1981.
- Лемеза Н., Шуханов Н. Малый практикум по низшим растениям. — Минск: Унiверсiтэцкае, 1994.
- Малахов В., Реунов А. Эволюция строения сперматозоидов у беспозвоночных // Успехи современной биологии. — Т. 113, вып. 1. — Стр. 3.
- Маргелис Л. Роль симбиоза в эволюции клетки. — М.: Мир, 1983.
- Медведев Н. Беседы по биологии пола. — Минск: 1976.
- Мирабдуллаев И. О происхождении и положении грибов в системе органического мира // Успехи современной биологии. — Т. 114, вып. 1.
- Мирабдуллаев И. Проблема происхождения эукариот // Успехи современной биологии. — Т. 107, вып. 3.
- Мирабдуллаев И. Проблемы классификации живого на уровне царств // Журнал общей биологии. — Т. 50, №6.
- Мирабдуллаев И. Рибосомы, кристы и филогения низших эукариот // Издание АН СССР, серия Биология. — 1989. — №5.
- Молчан И. Космизация курса общей биологии в системе опережающего образования и науки: прогнозы Живой Этики. // Три ключа (педагогический вестник). — 1998, вып. 2. — Стр. 36-51.
- Московский А. Существует ли научная альтернатива дарвинской концепции эволюции? // Три ключа (педагогический вестник). — 1999, вып. 3. — Стр. 49-54.
- Назаров В. Учение о макроэволюции. — М.: Наука, 1991.
- Наумов Н., Карташёв Н. Зоология позвоночных — в 2-х т. — М.: ВШ, 1979.
- Николаев Г. Магнитное поле Земли слабеет. Опасны ли последствия этого? // Наука и жизнь. — 1991, №1. — Стр. 44-50.
- Опарин А. Возникновение жизни на Земле. — М.: 57.
- Опарин А. Возникновение и начальное развитие жизни. — М.: Медицина, 1966.
- Павлинов И. Кладистический анализ. — М.: МГУ, 1990.
- Петров Ю. Основные этапы эволюции циклов развития у водорослей // Сб. «Проблемы филогении низших растений». — М.: Наука, 1974.
- Самовар А. Происхождение полового процесса и эукариот — взаимосвязанные проблемы? // Журнал эволюционной биохимии и физиологии. — №28. — С. 91.
- Северцев А. Направленность эволюции. — М.: МГУ, 1990.
- Симпсон Дж. Темпы и формы эволюции. — М.: ИИЛ, 1948.
- Стеббинс Дж., Айала Ф. Эволюция дарвинизма // В мире науки. — №8, 1984. — С. 26-35.
- Тарасов Денис. О неизбежности самозарождения жизни
во Вселенной и на Земле. —
http://lib.atheo-club.ru/index.php?action=show_article&a_id=13 - Тихоненко Т. Роль вирусов в обмене генетической информацией. — М.: Знание, 1980.
- Хесин Р. Непостоянство генома. — М.: Наука, 1984.
- Черникова И. Глобальный эволюционизм. — Томск: ТУ, 1987.
- Четвериков С. О некоторых моментах эволюционного процесса с точки зрения современной генетики (1926) // Классики современной генетики. — Л.: Наука, 68. — С. 133-170.
- Шкловский И. Вселенная, жизнь, разум. — М.: Наука, 1973.
- Эволюция генома. — М.: 1986.